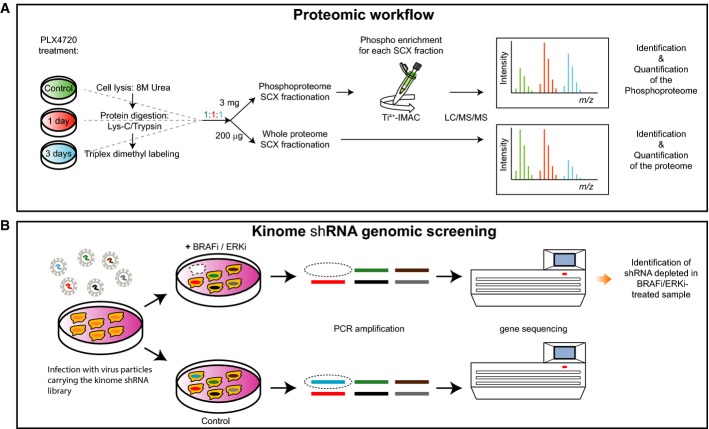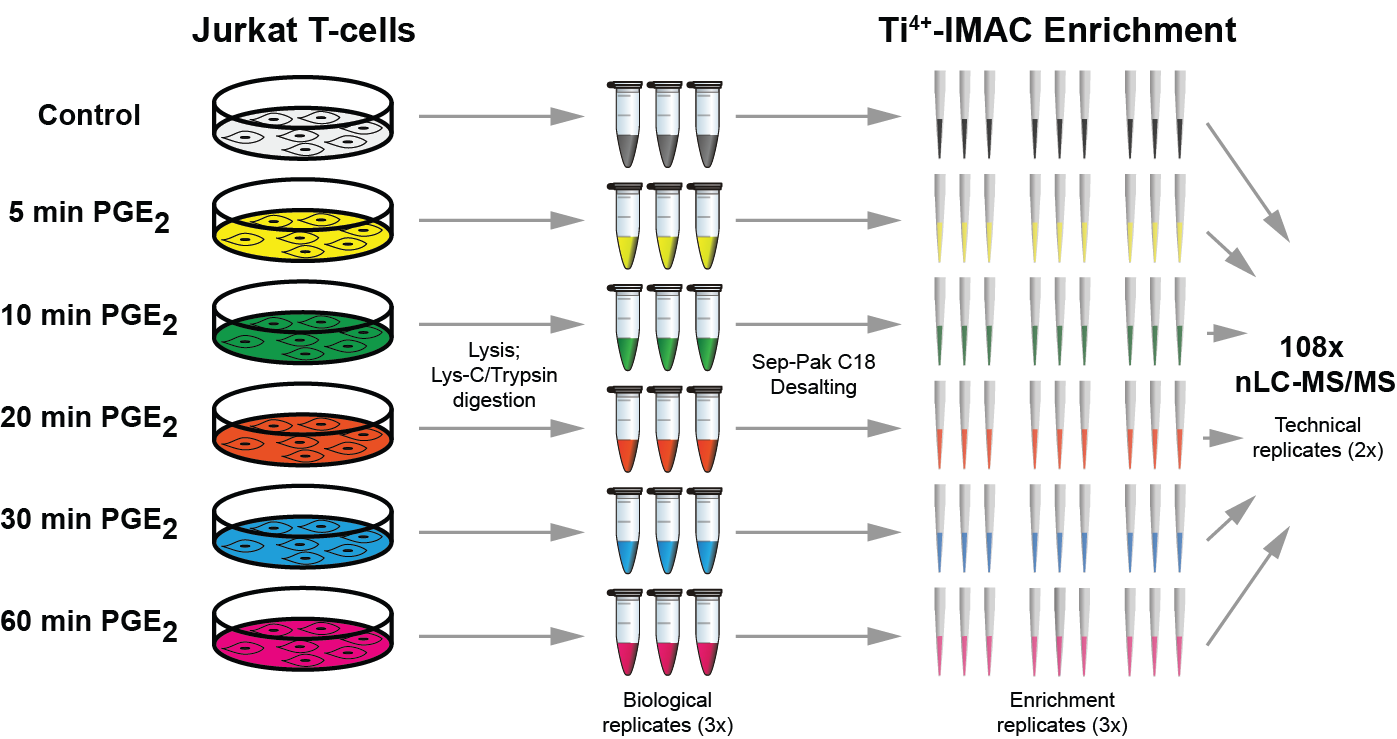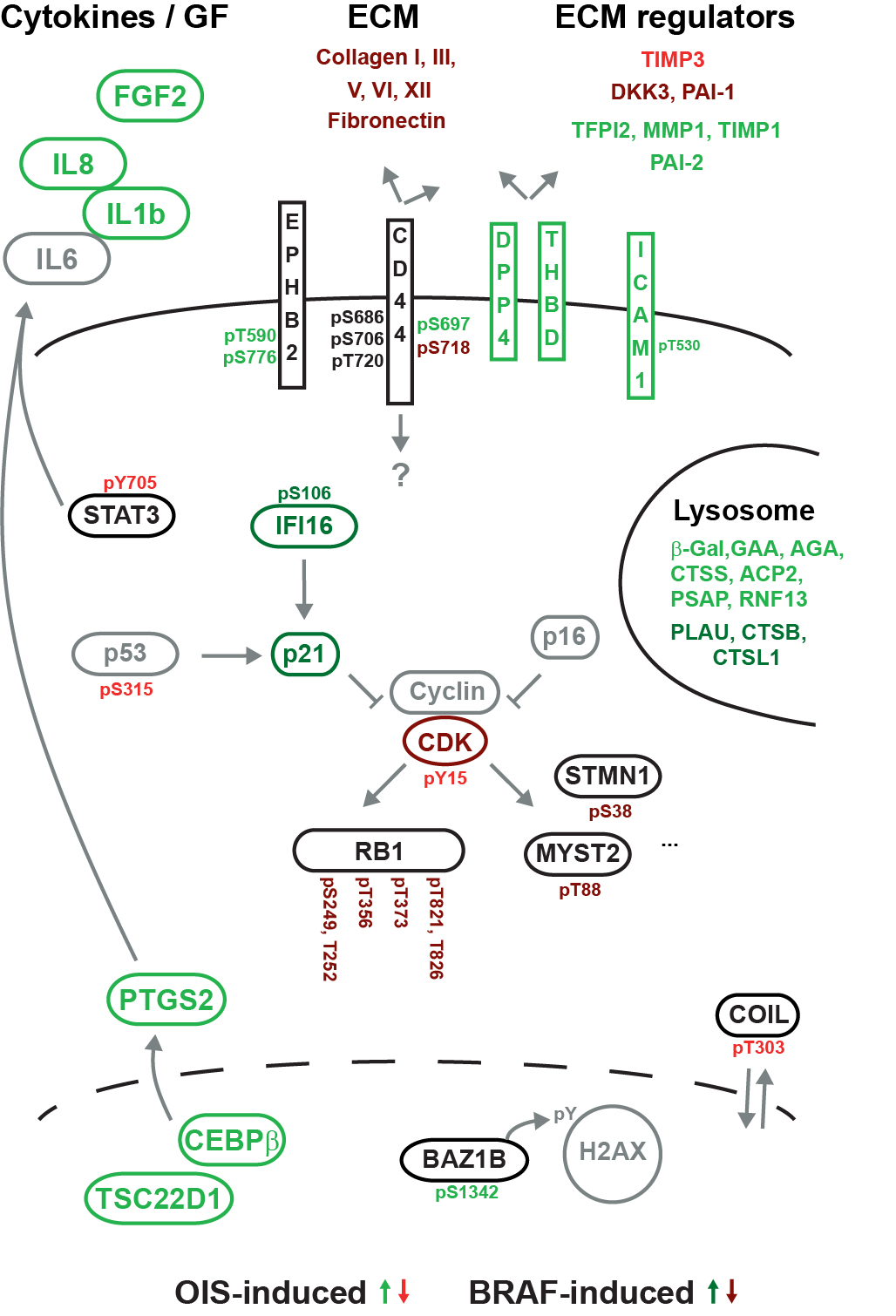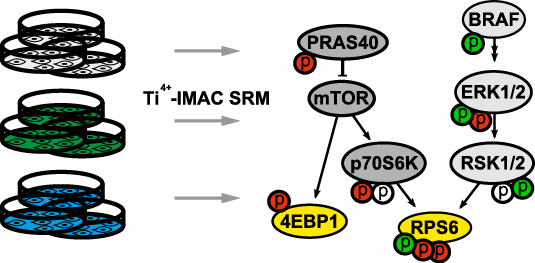
We report a straightforward strategy to comprehensively monitor signal transduction pathway dynamics in mammalian systems. Combining targeted quantitative proteomics with highly selective phosphopeptide enrichment, we monitor, with great sensitivity, phosphorylation dynamics of the PI3K-mTOR and MAPK signaling networks. Our approach consists of a single enrichment step followed by a single targeted proteomics experiment, circumventing the need for labeling and immune purification while enabling analysis of selected phosphorylation nodes throughout signaling pathways. The need for such a comprehensive pathway analysis is illustrated by highlighting previously uncharacterized phosphorylation changes in oncogene-induced senescence, associated with diverse biological phenotypes and pharmacological intervention of the PI3K-mTOR pathway.
de Graaf EL, Kaplon J, Mohammed S, Vereijken LA, Duarte DP, Redondo Gallego L, Heck AJ, Peeper DS, Altelaar AF.
J Proteome Res. 2015 Jul 2;14(7):2906-14.